1. Introduction
1.1. Background
1.1. Background
- When assessing the impact of a proposed offshore wind farm, it is crucial to determine the impact that such development will have on breeding seabird populations. Seabirds nest in colonies of variable sizes around the United Kingdom (UK) coastline (Burnell et al., 2023) and most species have large foraging ranges at sea (Woodward et al., 2019). Establishing the connectivity between marine renewable sites and colonies, which are often protected as Special Protected Areas (SPAs), is a key element of the assessment of impact. A theoretical approach has been developed by NatureScot (NatureScot, 2018) to determine the proportion of birds from SPA sites which use proposed development areas in the breeding season. In the non-breeding period, the standard approach to apportioning utilises the information presented in Furness (2015). These approaches allow the user to calculate apportioning values which are then used to ‘apportion’ the impact of a marine renewable project to multiple SPAs.
- This technical report presents the apportioning method and the seasonal apportioning values applicable to the Ossian Array (hereafter referred to as the “Array”) for SPAs that support qualifying species for which connectivity has been identified as part of the likely significant effect (LSE2) Screening Report (Part 1, appendix 1A; Ossian OWFL, 2023) (SPAs shown in Figure 1.1 Open ▸ ).
- Information and results from the following technical reports of the Array EIA Report (Ossian OWFL, 2024) were used to feed into the apportioning assessment:
- volume 3, appendix 11.1: Offshore Ornithology Baseline Characterisation Technical Report;
- volume 3, appendix 11.2: Offshore Ornithology Collision Risk Modelling (CRM) Technical Report; and
- volume 3, appendix 11.3: Offshore Ornithology Displacement Technical Report.
- The resulting apportioning values are presented for each protected site and will be used in the Report to Inform Appropriate Assessment (RIAA) to support the assessment of potential for an Adverse Effect on Integrity (AEOI) (Part 3).
Figure 1.1: SPAs Included in the Apportioning Assessment in Relation to the Offshore Array Area
2. Methodology
2. Methodology
2.1. Approach
2.1. Approach
- Apportioning undertaken for the breeding season is based on the NatureScot ‘theoretical approach’ method for the breeding season (NatureScot, 2018). Apportioning for the non-breeding period (i.e. post-breeding and pre-breeding seasons and in winter) generally utilises the Biologically Defined Minimum Population Scales (BDMPS) approach developed by Furness (2015). Seasonal definitions are set out in volume 3, appendix 11.1 of the Array EIA Report.
- In Scotland BDMPS is applied to all species with one exception. For guillemot Uria aalge NatureScot (2023a) advises that the mean-maximum foraging range + one standard deviation (SD) (i.e. the mean average of the maximum foraging trips recorded) should be used instead to determine connectivity to SPAs. As the Array lies beyond this distance (section 2.4) apportioning during the non-breeding period is not required under the NatureScot approach. However, Natural England in their Array EIA Scoping Report consultee response advised the non-breeding season for guillemot of Flamborough and Filey Coast SPA should be based on the BDMPS apportioning approach (Furness, 2015) (refer to the Array EIA Report volume 2, chapter 11, section 11.5; volume 2, appendix 6.2). This approach has been included within the apportioning undertaken. The advice from NatureScot (2023a) is based on tracking data of movements in the non-breeding season, and therefore the inclusion of Flamborough and Filey Coast SPA is considered to be over precautionary. Therefore, whilst this approach is included to meet Natural England’s request, it should be noted that the main assessment in the RIAA uses the approach recommended by NatureScot.
- For four species (shag Gulosus aristotelis, black-legged kittiwake Rissa tridactyla (hereafter referred to as kittiwake), guillemot and razorbill Alca torda), the Marine Scotland Apportioning Tool (Butler et al., 2020) is available to attribute seabirds to their breeding colonies, the use of which relies on Seabird 2000 (1998 to 2002) colony count data. These colony count data have recently been updated following the fourth UK breeding seabird census, Seabirds Count (JNCC, 2023), which focuses on the period 2015 to 2021, but has yet to be incorporated into the Marine Scotland Apportioning Tool. It is the view of NIRAS that to rely on an apportioning tool that requires the ‘old’ colony count data (based on the Seabird 2000 count) would produce unrealistic outputs. This is because in the intervening period, significant changes in colony size and, importantly, proportionate spread of the Scottish population between colonies has occurred (and as evidenced by scrutiny of the latest seabird census data). Therefore, the Marine Scotland Apportioning Tool has not been applied, with apportioning following the theoretical approach for all species for the breeding season as previously described. This proposed approach was shared with NatureScot during consultation correspondence in November 2023. Feedback on this point was not provided by NatureScot, but no objections or concerns were raised.
2.2. Identification of Species
2.2. Identification of Species
- Table 2.1 Open ▸ identifies the designated sites and associated features for which a LSE2 has been identified and therefore where apportioning values are required to apportion impacts from the Array. Although this table focuses on SPA populations, consideration has been given to all breeding colonies within the relevant foraging range of a species present within the Array survey area (as described in section 2.3, the approach to breeding season apportionment requires consideration of all colonies within foraging range, including those outside SPAs). Apportioning values for non-SPA colonies is provided in annex A.
Table 2.1: SPAs and Associated Qualifying Features for Which Apportioning is Required for the Array
2.3. Apportioning of Potential Impacts During the Breeding Season
2.3. Apportioning of Potential Impacts During the Breeding Season
- In the breeding season, a population of birds in a given sea area is likely to comprise of breeding adult birds from breeding colonies, immature birds (i.e. birds that have not yet reached breeding age) and non-breeding birds (i.e. birds that have reached breeding age but have not yet started breeding or are skipping a breeding season (sabbatical birds)). The proportion of each group must be estimated to allow the proportion of breeding birds to be identified, as it is this group that is relevant for Habitats Regulations Appraisal (HRA).
2.3.1. Breeding Adult Birds
2.3.1. Breeding Adult Birds
- The method followed NatureScot’s ‘Interim Guidance on Apportioning Impacts from Marine Renewable Developments to Breeding Seabird Populations in Special Protection Areas’ (NatureScot, 2018).
- To identify those breeding colonies (designated sites and other) for which there may be connectivity between breeding birds and the Array, the recommended foraging range given by NatureScot (2023b) has been used. In most cases, this is the mean-maximum foraging range + one SD as published by Woodward et al., (2019). However, a different foraging range, often informed by site-specific information, is recommended for certain species or protected sites. Guillemot is one such species, for which NatureScot advises use of a different foraging range under certain circumstances. For guillemot, NatureScot advise the use of the mean-maximum foraging range plus one standard for all Northern Isles SPAs (i.e. 153.7 km), and to use the mean-maximum foraging range plus one standard deviation calculated excluding data obtained from birds from Fair Isle for all designated sites south of the Pentland Firth (i.e. 95.2 km) (NatureScot 2023b).
- It should be noted that the Ossian Array Screening Report (Ossian OWFL, 2023) was drafted prior to the publication of NatureScot’s guidance and therefore used the mean-maximum foraging range + one SD as published by Woodward et al., (2019) for all species and sites. It should also be noted that the LSE2 Screening Report (Ossian OWFL, 2023) considered the shortest distance from the Array boundary to SPA boundaries when determining connectivity, while this report follows NatureScot (2018) in measuring distances from the centre of the Array to the centre of the colony. Therefore, the sites and species listed in this appendix will differ slightly from those identified in the LSE2 Screening Report (Ossian OWFL, 2023). Further details on changes since the LSE2 Screening Report (Ossian OWFL, 2023) are given in the main RIAA document (Parts 1 and 3).
- Following identification of breeding colonies with potential connectivity, three colony-specific weighting factors have been applied to each colony:
- colony size (with consistent count unit used between colonies for a species e.g. individuals, breeding pairs or apparently occupied sites);
- distance of colony from the development site; and
- sea area (the area extent of the open sea within the foraging range of the relevant species).
- Large colonies will contribute more individuals to the number of seabirds found at the Array, all other factors being equal. To account for this, a weighting factor based on colony size has been derived. Colony sizes for all species have been extracted directly from the recently published Seabirds Count (JNCC, 2023). The Seabirds Count is the fourth UK breeding seabird census, following on from the Seabird 2000 census. The Seabirds Count comprehensively surveyed seabird colonies, with surveys undertaken from 2015 to 2021[3]. As such, the Seabirds Count data represent the best available data on breeding seabird colony sizes (JNCC, 2023).
- Weighting by distance from the colony has been calculated using the measured distance between the geometric centre of the Array to the colony, using a by-sea route, as recommended by NatureScot (2018). The location of colonies was based on the coordinates given in Burnell et al. (2023). As birds move further away from a colony, density will decrease by a factor proportional to 1/distance2 as area increases proportionally by πr2. For the purposes of this assessment, a weighting factor based on 1/distance2 has been used as advised by NatureScot (2018).
- The available sea area for foraging has been measured by plotting a circle (defined by the species-specific foraging range around the colony) in ArcGIS and calculating the area of sea available to each seabird species. The fraction of the disc centred on the colony that is occupied by sea surface is then expressed as a decimal. As the density of birds is predicted to increase as the area of available foraging area decreases, this is used in the formula as 1/estimated area.
- The three weighting factors (weightings by colony size, distance from the colony and sea area) have been combined to produce an overall weighting for each colony. Each factor is given equal weight in the combined weighting. This calculation is provided below:
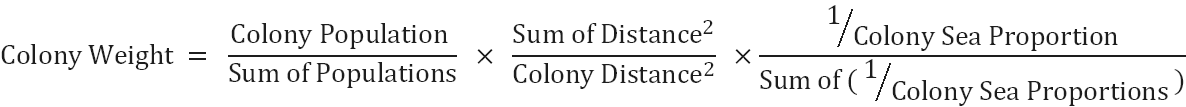
- The weighting was then used to calculate the proportion of birds attributed to each colony (“proportional weight of colony”) by calculating colony weight divided by sum of all colony weights.
- Where an SPA consists of multiple colonies, apportionment was carried out to each colony individually following the method described above. The total apportionment weight of the SPA population is then calculated as the sum of the weights of the individual colonies.
2.3.2. Immature Birds
2.3.2. Immature Birds
- A major part of any seabird population comprises immature birds. This is especially relevant for many of the species considered in this report, with some species not breeding until they reach six or more years of age (for example puffin, as taken from Horswill and Robinson, 2015). A proportion of immature birds return to natal waters during the breeding season, with the proportion of each immature age class increasing as individuals get closer to breeding age.
- To determine the proportion of immature birds present within the Array in the breeding season, data from the site-specific Digital Aerial Surveys (DAS) have been analysed ( Table 2.2 Open ▸ ). This approach can only be used for gannet, kittiwake and large gull species as they are easily distinguishable between immature and adult individuals. Identifying the age class of birds from other species, such as auks, using DAS is not feasible due to the similarity between immatures and adults. During site-specific surveys, no large gulls were assigned to any age class (volume 3, appendix 11.1, annex D of the Array EIA Report). Only those birds assigned to an age class have been included in the calculation in Table 2.2 Open ▸ . However, the number of birds for which an age class was not assigned is also provided. Those birds for which an age class can’t be determined are not included in the apportioning exercise. The data set out in Table 2.2 Open ▸ are used to inform the assessments undertaken in the RIAA.
Table 2.2: Number of Birds Assigned to Different Age Class Categories During Site-Specific Surveys of the Array Survey Area
- The identification of kittiwake age classes at sea is difficult and in most cases impossible (with the exception of first summer of younger birds). Whilst one year old kittiwakes can be easily identified due to differences in plumage, second and third year old birds, which have not yet reached the age of first breeding, cannot (Coulson, 2011; Olsen and Larsson, 2003). Therefore data on age class collected during surveys will potentially represent a considerable overestimate of the proportion of breeding adults present at the Array.
- To calculate an apportioning value for the breeding season in respect to the number of two and three year old kittiwakes at the Array, the analysis uses survival rates for immature kittiwake from Horswill and Robinson (2015) ( Table 2.3 Open ▸ ). The apportioned values will likely remain an under-estimate for the second and third year immatures as proportionately those cohorts show a much greater affinity for natal waters than first year birds.
Table 2.3: Estimated Breeding Season Contribution of Immature Kittiwake Predicted to be Present at the Array[4]
- For other species, birds cannot be readily aged from DAS imagery. Therefore, the proportion of adults present at the array in the breeding season is assumed to equal the proportion of adults within the population as a whole. The proportion of age classes is calculated from the demographic data in Horswill and Robinson (2015) assuming a stable age structure. This is presented in full in volume 3, appendix 11.1 of the Array EIA Report (Ossian WOFL, 2024), and the key values used for apportionment are repeated in Table 2.4 Open ▸ .
Table 2.4: Proportion of Age Classes of Other Species
- In the non-breeding season, the proportion of adults within the population does not need to be calculated or specified separately, as it is already factored into the Furness (2015) data used (see section 2.4).
2.3.3. Sabbaticals
2.3.3. Sabbaticals
- Every breeding season a proportion of adults skip breeding and take a ‘sabbatical’. The inclusion of sabbatical birds within the birds apportioned to a colony would likely lead to an overestimate of the effects to these species/populations (Marine Scotland 2017a; 2017b). This is because estimates of breeding colony population sizes used within the RIAA, do not include these sabbatical birds.
It is not possible to separate non-breeding adult birds from those that are breeding in a given sea area and therefore published estimates of sabbatical behaviour have been obtained ( Table 2.5 Open ▸ ). Consideration will be given in relevant assessments to the sabbatical values presented in
- Table 2.6 Open ▸ for each species, following the approach advised on other projects by the Scottish Ministers (2017a; 2017b, 2017c).
Table 2.5: Proportion of Sabbatical Birds to be Considered in the RIAA
Table 2.6: Proportion of Sabbatical Birds Used for RIAA Apportionment
2.4. Apportioning of Impacts During the Non-breeding Period
2.4. Apportioning of Impacts During the Non-breeding Period
2.4.1. Apportioning for all Species except Guillemot
2.4.1. Apportioning for all Species except Guillemot
- The calculation of apportioning values for non-breeding seasons (post-breeding, non-breeding and pre-breeding) has followed the approach advised by NatureScot (2023a). For species except for guillemot, the contribution of adult birds from an individual designated site, as estimated by Furness (2015), to the relevant BDMPS population for each species/season combination is divided by the total BDMPS population. The calculated value is the proportion of the BDMPS population represented by adult birds from the designated site under consideration. It should be noted that no updates have been made to the population data presented in Furness (2015) as selectively updating specific colonies would cause inconsistencies with the original dataset. For any designated site not named in Furness (2015), the proportion of birds present in the relevant BDMPS areas in each season has been taken from the nearest named designated site.
2.4.2. Apportioning for Guillemot
2.4.2. Apportioning for Guillemot
- As set out in paragraph 6, guillemot is the only species for which NatureScot (2023a) advises an alternative approach to the use of the BDMPS approach (Furness, 2015) when determining which colony SPAs have connectivity with the Array during the non-breeding season.
- As the Array lies beyond the foraging range parameter values (as set out in section 2.3.1), no connectivity with colony SPAs have been identified and therefore no apportioning has been undertaken for guillemot following the NatureScot advised approach. However, Natural England in their screening responses to Ossian, advised of their expectation for the apportioning undertaken for the Array during the non-breeding season for guillemot from Flamborough and Filey Coast SPA, to be based on the BDMPS apportioning approach (Furness, 2015). Therefore, apportioning has been undertaken using the BDMPS apportioning approach as described in the previous paragraph for all other species, but presenting only the result for Flamborough and Filey SPA.
3. Results
3. Results
- The results of the apportioning exercise are presented in sections 3.1 to 3.5 below.
- The proportions, weights and apportioning values in the tables in sections 3.1 to 3.5 are values that are scored out of one (i.e. 1.0000 equates to 100%, 0.1000 equates to 10% and 0.0100 equates to 1%).
3.1. Kittiwake
3.1. Kittiwake
- Apportioning values in the breeding season for all colonies within foraging range of the Array is presented in Table 3.1 Open ▸ , with SPA colonies shown in Figure 3.1 Open ▸ . Apportioning values for non-SPA colonies is presented in annex A. Apportioning values for use in the post-breeding and pre-breeding seasons are presented in Table 3.2 Open ▸ .
Figure 3.1 SPAs Included in the Apportioning Assessment for Kittiwake
Table 3.1: Apportioning Values for Kittiwake in the Breeding Season for SPAs within Foraging Range
Table 3.2: Post-breeding and Pre-breeding Season Apportioning Values for Kittiwake
3.2. Guillemot
3.2. Guillemot
- Apportioning value for guillemot at Flamborough and Filey Coast SPA in the non-breeding season is presented in Table 3.3 Open ▸ . The location of the SPA colony is shown in Figure 3.2 Open ▸ .
Table 3.3: Apportioning Values for Guillemot in the Non-breeding Season for Flamborough and Filey Coast SPA
Figure 3.2 SPAs Included in the Apportioning Assessment for Guillemot
3.3. Razorbill
3.3. Razorbill
- Apportioning values in the breeding season for all colonies within foraging range of the Array is presented in Table 3.4 Open ▸ , with SPA colonies shown in Figure 3.3 Open ▸ . Apportioning values for non-SPA colonies is presented in annex A. Apportioning values for use in the post-breeding, winter and pre-breeding seasons are presented Table 3.5 Open ▸ .
Figure 3.3 SPAs Included in the Apportioning Assessment for Razorbill
Table 3.4: Apportioning Values for Razorbill in the Breeding Season for SPAs Within Foraging Range
Table 3.5: Calculation of Post-breeding, Winter, and Pre-breeding Season Apportioning Values for Razorbill
3.4. Puffin
3.4. Puffin
- Apportioning values in the breeding season, for all colonies within foraging range of the Array, is presented in Table 3.6 Open ▸ , with SPA colonies shown in Figure 3.4 Open ▸ . Apportioning values for non-SPA colonies is presented in annex A. Apportioning values were calculated for the breeding season only for puffin.
Table 3.6: Apportioning Values for Puffin in the Breeding Season for SPAs Within Foraging Range
Figure 3.4 SPAs Included in the Apportioning Assessment for Puffin
3.5. Gannet
3.5. Gannet
- Apportioning values in the breeding season, for all colonies within foraging range of the Array, is presented in Table 3.7 Open ▸ , with SPA colonies shown in Figure 3.5 Open ▸ . Apportioning values for non-SPA colonies is presented in annex A. Apportioning values for use in the post-breeding and pre-breeding seasons are presented in Table 3.8 Open ▸ .
Table 3.7: Apportioning Values for Gannet in the Breeding Season for SPAs Within Foraging Range of the Array
Figure 3.5 SPAs Included in the Apportioning Assessment for Gannet
Table 3.8: Post-breeding and Pre-breeding Season Apportioning Values for Gannet
4. Discussion
4. Discussion
- This technical report uses NatureScot’s theoretical approach to apportioning impacts to breeding seabirds in SPAs and its consideration of immature birds.
- For kittiwake, in the breeding season, most birds are apportioned to Fowlsheugh SPA, Buchan Ness to Collieston Coast SPA or Flamborough and Filey Coast SPA. In the migratory seasons, Flamborough and Filey Coast SPA is again relatively important, and additionally East Caithness Cliffs SPA is relatively important – although it should also be noted that in the migratory seasons, birds come from a much wider range of SPAs and non-SPA colonies (including overseas colonies) and therefore the proportion attributed to SPAs with an LSE2 identified are small.
- For razorbill, only one SPA (Fowlsheugh SPA) has been identified as having potential LSE2.
- For puffin, in the breeding season most birds are apportioned to either the Farne Islands SPA or the Forth Islands SPA. In the non-breeding season, puffin are expected to disperse widely and apportionment is not carried out.
- For gannet, in the breeding season the majority of birds are apportioned to the Forth Islands SPA (which includes the UK’s largest breeding gannet colony at Bass Rock). In the non-breeding season, of the SPAs screened in, the Forth Islands SPA still has the highest apportionment value, although it should also be noted that in the migratory seasons, birds come from a much wider range of SPAs and non-SPA colonies (including overseas colonies) and therefore the proportion attributed to the Forth Islands SPA or any other SPAs with an LSE2 identified are small.
- For gannet, it should be noted that other projects (SSE Renewables, 2022; Seagreen Wind Energy Ltd, 2019; MacArthur Green, 2015) have considered an alternative approach to apportioning gannets in the migratory seasons to SPA populations. This alternative approach uses tracking data to infer the number of birds that may pass through a windfarm on migration, based on the location of the wind farm compared to all North Sea breeding colonies, and the typical direction of migration in the post-breeding and pre-breeding migration seasons. Whilst this approach has not been applied, it should be noted that this approach would be expected to result in fewer birds being apportioned to the Forth Islands SPA and the Flamborough and Filey Coast SPA, especially in the post-breeding season, due to the majority of birds migrating southwards towards the English Channel and therefore being less likely to interact with the Array (MacArthur Green, 2015).
- Using the approach recommended by NatureScot, no guillemot affected by the Array are expected to be birds associated with SPA colonies and therefore no apportionment is required in any season. However, following Natural England’s request, apportionment of guillemot to Flamborough and Filey Coast SPA during the non-breeding season has also been carried out using the method preferred by Natural England, and this is provided without prejudice for information only.
- The results include several assumptions that may lead to an over-estimate of the proportion of breeding adult birds present in a given area. These assumptions are listed below.
- breeding adult birds:
– the assumption that birds are evenly distributed at sea, with this being extremely unlikely due to the known patchy distribution of prey species and information gained from tracking studies;
– the assumption that seabird colonies are independent of one another;
– the assumption that larger foraging ranges will occur at larger breeding colonies due to competition and prey depletion closer to the colony (Elliott et al., 2009); and
– the use of mean-maximum plus one standard deviation foraging ranges.
- immature birds:
– limited information is available on the proportion of immature birds that return to natal waters and the distribution of immature birds within natal waters.
- Consideration has been given in the RIAA to these assumptions/limitations, including where available site-specific tracking studies, and what effect they may have on the overall magnitude of any impacts (Ossian OWFL, 2024). Further information on foraging range and immature proportions is provided in section 4.1 and 4.2 below.
4.1. Foraging Range
4.1. Foraging Range
- NatureScot (2023b) typically recommends the use of the mean-maximum foraging range plus one standard deviation for each species considered.
- The use of a mean-maximum foraging range plus one standard deviation represents a highly precautionary approach regardless of its application as, although it ensures, from a HRA screening perspective, that no SPAs are erroneously omitted from the RIAA, the likelihood of an LSE2 occurring as a result of any project beyond mean-maximum foraging range is considered highly unlikely. A mean-maximum foraging range already represents the average of the maximum foraging ranges exhibited by birds across multiple studies. A standard deviation of a mean value represents the amount by which individual values differ from the mean value. It is an expression of confidence in the mean value and should not be applied as an absolute value as in the application of foraging ranges for screening. This is particularly so when the average value is already an average of maximum values from multiple studies which may not reflect the true foraging behaviour of all individuals from a colony.
- It is considered that the application of a mean-maximum foraging range plus one SD for apportioning purposes is likely to be over precautionary in terms of including distant SPAs for assessment, for which the true impact is likely to be negligible or zero. It also leads to impacts being diluted across more SPAs and therefore potentially underestimates impacts on nearby colonies. However, this is the recommended approach and therefore has been applied in this apportioning assessment.
4.2. Immature proportions
4.2. Immature proportions
- Although any population of breeding seabirds has an immature component associated with it, the spatial distribution of the immature component is often very different to the breeding adult component, especially in the breeding season. For many seabird species, immature birds gradually begin to return to natal waters in the breeding season as they get nearer to breeding age. The proportion of older immature age classes in natal waters is therefore higher than the proportion of younger immature age classes. In addition, the distribution of immature birds in natal waters may be dictated by proximity to breeding colonies either because birds are prospecting for breeding sites or due to competition with breeding adult birds. Where all immature classes of a species can be reliably identified during baseline surveys, this is less of an issue. However, for species for which only some age classes can be identified during baseline surveys (i.e. the birds obtain an adult type plumage before they are physically old enough to breed), resulting immature proportions represent an under-estimate.
- In the context of this apportioning appendix this is relevant to kittiwake. Whilst one year old kittiwakes can be easily identified due to differences in plumage, second and third year old birds, which have not yet reached the age of first breeding, cannot (Coulson, 2011; Olsen and Larsson, 2003). Therefore data on age class collected during surveys will potentially represent a considerable overestimate of the proportion of breeding adults present in a given sea area. As set out in section 2.3.2, the proportion of second and third year immatures (which show a much greater affinity for natal waters than first year birds) will be under-estimated.
- As detailed in section 2.3, an approach has been applied which aims to address the underestimation. Whilst maintaining the proportion represented of each year class of immatures at the Array, mortality reduces the absolute number of birds present from each successive year class of kittiwake. In calculating the number of two and three year old kittiwakes at the Array, the analysis uses survival rates of each immature age class of kittiwake that follows the rate provided in Horswill and Robinson (2015). This approach is considered precautionary for the following reasons:
- it is known that older immature age classes that are not identifiable during baseline surveys will be present at the Array; and
- a smaller proportion of one year old birds are likely to be present in natal waters with a much greater proportion of older age classes of immature birds showing affinity with natal waters and therefore the proportions of older age classes is likely underestimated when applying the approach.
- The identification of immature age classes of gannets during baseline surveys is far easier than for kittiwakes and the immature proportions calculated for these species are therefore considered to be more representative.
- The identification of immature age classes of auk species is not possible from baseline surveys (with the exception of juvenile birds in the post-breeding season) and, where necessary, other sources will be relied upon within the RIAA.
5. References
5. References
Burnell, D., Perkins, A. J., Newton, S.F., Bolton, M., Tierney, T.D., Dunn, T.E. (2023). Seabirds Count: a census of breeding seabirds in Britain and Ireland (2015-2021). Lynx Nature Books, Barcelona.
Butler, A., Carroll, M., Searle, K., Bolton, M., Waggitt, J., Evans, P., Rehfisch, M., Goddard, B., Brewer, M., Burthe, S. and Daunt, F. 2020. Attributing seabirds at sea to appropriate breeding colonies and populations (CR/2015/18). Scottish Marine and Freshwater Science Vol 11 No 8, 140pp. DOI: 10.7489/2006-1.
Coulson, J.C. (2011). The Kittiwake. T and AD Poyser, London, UK. ISBN: 978-1-4081-0966-3.
Coulson, J.C. (2019). Gulls. HarperCollins, London, UK. ISBN: 978-0-00-820142-5.
East Anglia THREE Ltd. (2015). East Anglia THREE Information for the Habitats Regulations Assessment. Document Reference 5.4.
Elliott K.H., Woo K.J., Gaston A. J., Benvenuti S., Dall’Antonia L., and Davoren G. K. (2009) Central-place Foraging in an Arctic Seabird Provides Evidence for Storer- Ashmole’s Halo. The Auk ,126, 613−625.
Forewind (2013). Dogger Bank Creyke Beck Information for Appropriate Assessment Report. F-OFC-RP-002 Issue 11. Application Reference: 5.2.
Furness, R.W. (2015). Non-breeding season populations of seabirds in UK waters: Population sizes for Biologically Defined Minimum Population Scales (BDMPS). Natural England Commissioned Reports, No. 164.
Horswill, C. and Robinson, R.A. (2015). Review of Seabird Demographic Rates and Density Dependence. JNCC, Peterborough.
JNCC (2023). Seabird Censuses. Available at: https://jncc.gov.uk/our-work/seabird-censuses/. Accessed on: 05 February 2024.
MacArthur Green (2015). East Anglia THREE. Information for Habitats Regulations Assessment. Appendix 3: Apportioning of the Flamborough Head and Filey Coast pSPA Gannet Population among North Sea Offshore Windfarms.
Marine Scotland (2017a). Marine Scotland - Licensing Operations Team Scoping Opinion. Addendum: Ornithology. Scoping Opinion for Inch Cape Offshore Windfarm – Revised Design Parameters – Ornithology. 10 August 2017.
Marine Scotland (2017b). Marine Scotland - Licensing Operations Team Scoping Opinion. Addendum: Ornithology. Scoping Opinion for Moray East Offshore Windfarm – Alternative Design Parameters – Ornithology. 16 June 2017.
Marine Scotland (2017c). Marine Scotland - Licensing Operations Team Scoping Opinion. Scoping Opinion For The Proposed Section 36 Consent And Associated Marine Licence Application For The Revised Seagreen Phase 1 Offshore Project. 15 September 2017.
NatureScot (2018). Interim Guidance on apportioning impacts from marine renewable developments to breeding seabird populations in SPAs.
NatureScot (2023a). Guidance Note 4: Guidance to Support Offshore Wind Applications: Ornithology - Determining Connectivity of Marine Birds with Marine Special Protection Areas and Breeding Seabirds from Colony SPAs in the Non-Breeding Season. Available at: https://www.nature.scot/doc/guidance-note-4-guidance-support-offshore-wind-applications-ornithology-determining-connectivity. Accessed on: 01 March 2024.
NatureScot (2023b). Guidance Note 3: Guidance to support Offshore Wind applications: Marine Birds - Identifying theoretical connectivity with breeding site Special Protection Areas using breeding season foraging ranges. Available at: https://www.nature.scot/doc/guidance-note-3-guidance-support-offshore-wind-applications-marine-birds-identifying-theoretical. Accessed on: 01 March 2024.
Olsen, K.M. and Larsson, H. (2003). Gulls of Europe, Asia and North America. Christopher Helm, London.
Ossian Offshore Wind Farm Ltd (2023). Ossian Array Habitats Regulations Appraisal Stage 1: Likely Significant Effects Screening Report. Available at: https://marine.gov.scot/node/23666. Accessed on: 22 February 2024.
Ossian OWFL (2024). Array Environmental Impact Assessment.
Seagreen Wind Energy Ltd. (2019). May 2019 Addendum: Part 2 Ornithology Habitats Regulations Appraisal. Available at https://www.seagreenwindenergy.com/_files/ugd/fe5128_5d0c70d6629e4bf8b87c20e8c8df41f8.pdf. Accessed April 2024
SSE Renewables (2022). Berwick Bank Wind Farm Offshore Environmental Impact Assessment Appendix 11.5: Ornithology Apportioning Technical Report. Available at https://berwickbank-eia.com/offshore-eia/vol3-ap1105-vol3-apportioning/part-1.html. Accessed April 2024.
SMart Wind (2015). Hornsea Offshore Wind Farm Project Two Habitats Regulations Assessment. Report to Inform the Habitats Regulations Assessment. Document Reference 12.6. APFP Regulation 5(2)(g). January 2015. SMart Wind Limited, London.
Woodward, I., Thaxter, C.B., Owen, E. and Cook, A.S.C.P. (2019). Desk-based revision of seabird foraging ranges used for HRA screening. Available at: http://www.marinedataexchange.co.uk/ Accessed on: 22 February 2024
Annex A: Breeding Season Apportioning Values for Non-SPA Colonies
Annex A: Breeding Season Apportioning Values for Non-SPA Colonies
- Please find Annex A attached to this document separately.
[1] Where a SPA consists of multiple colonies, the distance given is the average distance.
[2] Included in the non-breeding season following Natural England’s scoping response; see section 2.1 for explanation.
[3] The data from the Seabirds Count are assumed to be unaffected by the HPAI outbreak, which did not significantly impact breeding seabird colonies until 2022.
[4] Using immature proportions as calculated from survival rates and numbers of one year old birds recorded on baseline survey transects covering the Array.
[5] Apparently Occupied Nests (bird census) (AON)
[6] Where a colony consists of multiple subsites average values are provided for distance to the Offshore Array Area and proportion of foraging range at sea.
[7] Number of individuals counted (bird census) (IND)
[8] Apparently Occupied Burrow (AOB)
[9] Number of individuals counted (bird census) (IND)