Socio-economics
- For socio-economics, the following potential impacts have been considered within the inter-related effects assessment:
- employment and GVA impacts associated with the construction, operation and maintenance and decommissioning of the Array; and
- demographic changes and demand for housing and other services.
- Table 20.13 Open ▸ lists the inter-related effects (Array lifetime effects) that are predicted to arise during the construction, operation and maintenance, and decommissioning phases of the Array and also the inter-related effects (receptor-led effects) that are predicted to arise for socio-economic receptors.
Table 20.13: Summary of Likely Significant Potential Inter-Related Effects for Socio-economics from Individual Effects Occurring across the Construction, Operation and Maintenance and Decommissioning Phases of the Array (Array Lifetime Effects) and from Multiple Effects Interacting Across all Phases (Receptor-led Effects)
Marine archaeology
- For marine archaeology, the following potential impacts have been considered within the inter-related effects assessment:
- sediment disturbance and deposition leading to indirect impacts on marine archaeology receptors; and
- alteration of sediment transport regimes.
- Direct damage to marine archaeology receptors has not been assessed as part of the inter-related effects assessment as there is no potential for direct damage to accumulate through the lifetime of the project or to interact, spatially and temporally, to create inter-related effects on a receptor.
- Table 20.14 Open ▸ lists the inter-related effects (Array lifetime effects) that are predicted to arise during the construction, operation and maintenance, and decommissioning phases of the Array and also the inter-related effects (receptor-led effects) that are predicted to arise for infrastructure and other users.
- Effects on marine archaeology do not have the potential to lead to secondary effects on other receptors.
Table 20.14: Summary of Likely Significant Potential Inter-Related Effects for Marine Archaeology from Individual Effects Occurring across the Construction, Operation and Maintenance and Decommissioning Phases of the Array (Array Lifetime Effects) and from Multiple Effects Interacting Across all Phases (Receptor-led Effects)
20.8.2. Cumulative Effects Assessment
- As mentioned in paragraph 25, the inter-related effects from other projects are considered in the cumulative effects sections of the relevant chapters (volume 2, chapters 7 to 20). However, the cumulative effects of the Proposed offshore export cable are considered in this section of this chapter.
- The Proposed offshore export cable is considered unlikely to have the potential to result in a cumulative impact with the Array, other than in very close proximity to the Array. This is due to the nature and scale of the proposed offshore export cable works which are likely to be restricted in nature and associated only with cable laying. The total footprint is not available at present.
- At the time of writing this Array EIA Report, there was no EIA Report available for the Proposed offshore export cable corridor(s), but the activities and footprints of disturbance associated with its site preparation and construction phase are expected to be similar to those of other cable laying projects e.g. Easter Green Link 2.
- Activities associated with the site preparation and construction phase for the Proposed offshore export cable corridor(s) are expected to be of an equal or lesser extent than those represented by the MDS for the Array alone for all relevant topics. The impacts of cable installation and seabed preparation are likely to be reversible. The cumulative magnitude of impact of the Array with the proposed offshore export cable is therefore not expected to represent a material additional impact to that defined for the assessment of the Array alone for any assessment presented in the volume 2, chapters 7 to 20. Within this phase of development of the Array, site preparation and construction activities are anticipated to occur intermittently. They will be spread out across the full allotted timeframe with only a small proportion of the MDS footprint for this impact being affected at any one time. There may be some spatial overlap between the Array and the Proposed offshore export cable corridor(s), given their proximity, however this is considered to be of very short duration and limited in extent.
- The cumulative impact is therefore predicted to be of local spatial extent, short term duration (between 2030 and 2038), intermittent, and of high reversibility. It is predicted that the impact will affect the receptors directly. The magnitude is therefore considered to be low and the sensitivity of the receptors is as set out in volume 2, chapters 7 to 20. The overall significance of assessment is therefore considered to be minor adverse significance, which is not significant in EIA terms.
20.9. Part Two: Ecosystem Based Effects Assessment
20.9.1. Overview
- An ecosystem is a community of living (biotic) organisms existing in conjunction with the non-living (abiotic) components of their environment. These biotic and abiotic components are linked together through nutrient cycles and energy flows (LibreTexts, 2022). In marine ecosystems biotic components include plankton, seaweed, benthic communities, fish, seabirds and marine mammals and abiotic components include air, salt water, seabed components and rock.
- Biodiversity is defined as the collection of genomes, species, and ecosystems occurring in a geographically defined region (National Research Council, Division on Earth, Life Studies, Commission on Geosciences, Resources and Committee on Biological Diversity in Marine Systems, 1995). The biodiversity within an ecosystem is the key indicator of the health of an ecosystem. A wider variety of species will cope better with external pressures than a limited number of species in large populations. Even if certain species are affected by climate change or human activities, the ecosystem as a whole may adapt and survive (European Commission, 2022).
- The purpose of this ecosystem-based assessment is to qualitatively assess the potential effects of the Array at the ecosystem level, to better understand how predator – prey relationships could be altered and how this could impact the functioning of the ecosystem. This is to address the advice raised by NatureScot during consultation that “increasingly there is a need to understand potential impacts holistically at a wider ecosystem scale in addition to the standard set of discrete individual receptor assessments. This assessment should focus on potential impacts across key trophic levels particularly in relation to the availability of prey species. This will enable a better understanding of the consequences (positive or negative) of any potential changes in prey distribution and abundance from the development of the wind farm on seabird and marine mammal (and other top predator) interests and what influence this may have on population level impacts”.
20.9.2. Ecosystem Baseline
- This section provides a summary of the abiotic and biotic components of the marine ecosystem relevant to the Array, considering the topic specific study areas used for assessment.
- The Array will be located within the site boundary, located off the east coast of Scotland, approximately 80 km south-east of Aberdeen from the nearest point, and comprising an area of approximately 859 km2. Across the site boundary, the maximum water depth was recorded at 88.7 m Lowest Astronomical Tide (LAT), and the shallowest area was recorded at 63.8 m LAT. The seabed across the site boundary is relatively flat with a gentle slope downwards in an approximately north-west to south-east direction (Ocean Infinity, 2022). The average water depth across the site boundary is 74.47 m below LAT.
- The seabed within the site boundary consists primarily of sand, with some areas of gravel and occasional diamicton (poorly sorted mixed sediments). Gravel areas are more frequent in the north-west, with occasional diamicton also observed in this area.
- The geological morphology within the site boundary is varied and includes the following features:
- megaripples;
- sand waves;
- boulders (primarily in the north-west);
- recent marine soft sediment deposits; and
- deep channel structures (down to 60 m) with sedimentary infill (south-eastern corner).
- The benthic communities within the site boundary are characterised by polychaetes (particularly bristleworm Spiophanes bombyx), dead man’s fingers Alcyonium digitatum, and various echinoderms and bryozoans (such as hornwrack Flustra foliacea). Biomass between grab sampling sites was varied, with six major phyla identified: Echinodermata, Mollusca, Annelida, Arthropoda, Cnidaria and Bryozoa. Echinoderms comprised the majority of the biomass within the grab samples (65%), which is largely due to the purple heart urchin Spatangus purpureus and sea potato Echinocardium cordatum occurring at several grab sampling sites. The phyletic composition was dominated by annelids, mainly sand mason worm Lanice conchilega and S. bombyx. The phyletic composition of sessile colonial fauna was dominated by cnidarians and bryozoans, with cnidarians representing the highest number of taxa and bryozoans the highest number of colonies. The closest designated site (located approximately 25 km west of the Array) is the Firth of Forth Banks Complex Marine Protected Area (MPA) which is designated for ocean quahog Arctica islandica, offshore subtidal sand and gravels, shelf banks and mounds, moraines representative of the Week Bankie Key Geodiversity Area (volume 2, chapter 8).
- Table 20.15 Open ▸ provides a summary of the seven main broad subtidal habitats present with the site boundary. Details of the grab sample locations (e.g. S018) can be found in volume 2, chapter 8.
Table 20.15: Broad Habitat Types and Species
20.9.3. The Marine Food Web
- Trophic levels describe the hierarchical levels which organisms occupy in the food web. Primary producers, such as phytoplankton and seaweed, form the lowest trophic levels in marine food webs. They are consumed by primary consumers (herbivores) such as zooplankton, some crustaceans (e.g. copepods) and molluscs (e.g. clams, snails, mussels). Secondary consumers (carnivores or omnivores) such as fish larvae, Atlantic herring Clupea harengus (hereafter referred to as “herring”) and lesser sandeel Ammodytes marinus, and some crustaceans (e.g. crabs, shrimp) feed on primary consumers and primary producers. These species support tertiary consumers (carnivores), including some fish species, and cephalopods (e.g. octopus and squid species). Seabirds, along with marine mammals, large marine fish and elasmobranchs (sharks, skates and rays), are the top predators of the natural marine food web. An example of a marine food web which illustrates the interactions between the different trophic levels is presented in Figure 20.1.
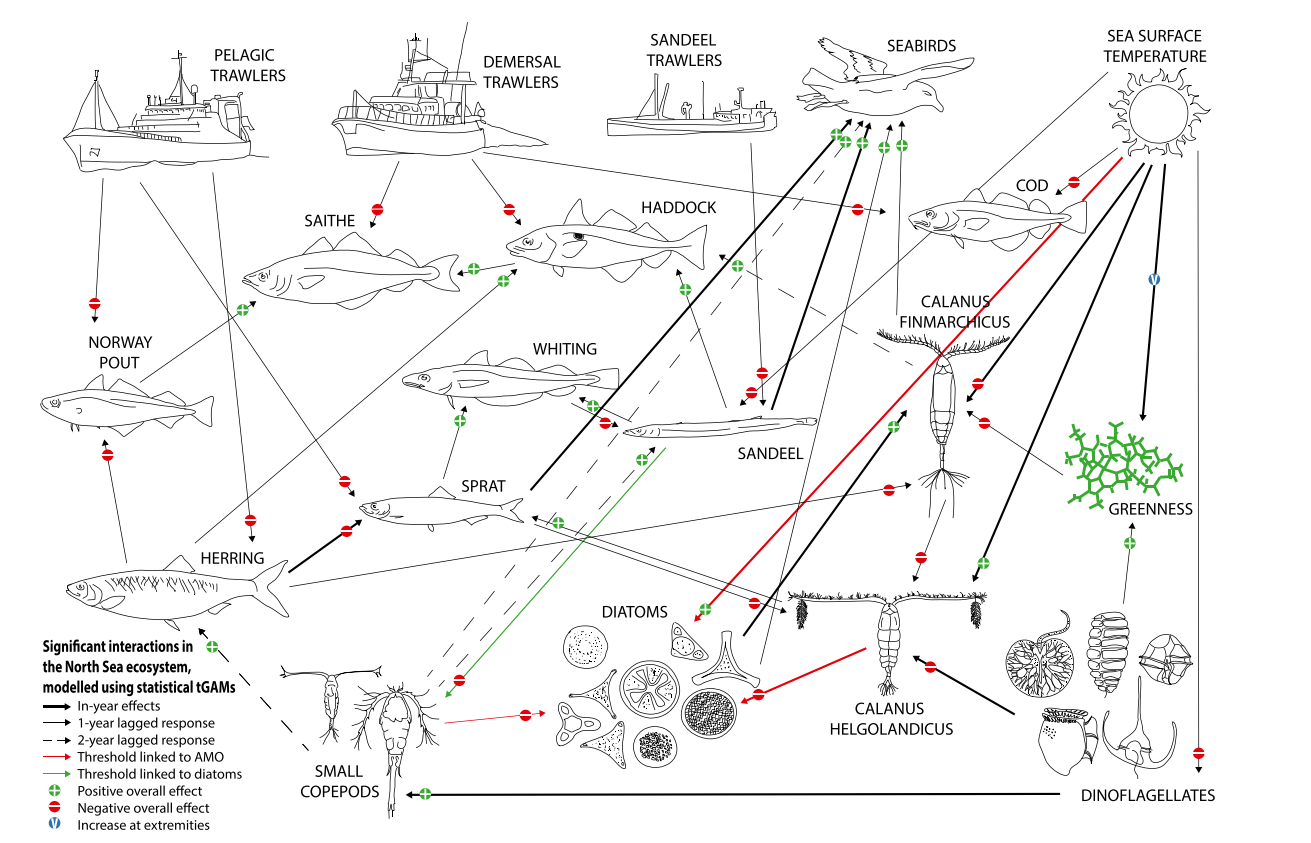
Figure 20.1: Significant Interactions Between Functional Groups and Drivers (from Lynam et al., 2017)
20.9.4. The Key Predator Species
- Volume 2, chapters 9, 10 and 11, provide details on the fish, marine mammals and seabirds which are most abundant in the associated topic study areas and are the receptors most likely to be impacted by activities associated with all phases of the Array. From information on these receptor groups it is possible to ascertain which fish, seabird and marine mammal species are likely to be key predators in the marine ecosystem in this part of the central North Sea and within the study areas outlined in section 20.3.
Piscivorous fish
- The key marine predatory fish likely to utilise the marine environment within the site boundary are cod, haddock Melanogrammus aeglefinus, whiting Merlangius merlangus, plaice Pleuronectes platessa, saithe Pollachius virens and European hake Merluccius poutassou (hereafter referred to as “hake”). Piscivorous fish primarily feed on other fish species with these diet of these species including small forage such as sandeel, juvenile whiting and juvenile haddock. Several elasmobranch species are also likely to be present whose diet includes small forage food such as tope shark Galeorhinus galeus, spurdog Squalus acanthias, common skate Dipturus batis and rays.
- The migration route of diadromous fish species which also feed on small forage fish, and which are likely to pass through the site boundary during their migration (volume 3, appendix 9.1) are Atlantic salmon Salmo salar, sea trout Salmo trutta, sea lamprey Petromyzon marinus, European eel Anguilla anguilla, allis shad Alosa alosa and twaite shad Allosa fallax.
- Table 20.16 Open ▸ lists the key predator species and the prey they feed on. This shows that although sandeel, herring, mackerel Scomber scombrus and European sprat Sprattus sprattus (hereafter referred to as “sprat”) are components of most of the key predators’ diets, other fish and benthic fauna are also important in the diet of marine predatory fish.
Table 20.16: Key Predatory Fish Species and their Prey
Marine mammals
- The key marine mammal species which are most abundant within and therefore have the potential to be impacted by the Array are:
- harbour porpoise;
- bottlenose dolphin;
- white-beaked dolphin Lagenorhynchus albirostris;
- minke whale Balaenoptera acutorostrata ;
- humpback whale Megaptera novaeangliae; and
- grey seal.
- These species correspond to the marine mammal IEF identified in volume 2, chapter 10. The sensitivity of marine mammals to prey availability within the site boundary will be affected by how important this area is to each species and how sensitive they are to prey availability. This is discussed further in section 20.9.10.
- A summary of the dietary preferences of key marine mammal species within the marine mammal study area is presented in Table 20.17 Open ▸ . Further details of the most appropriate density values for marine mammals that have then been taken forward in the assessment are detailed in volume 3, appendix 10.2.
Table 20.17: Diet and Abundance of Key Marine Mammal Species
Seabirds
- The key seabird species which are most abundant (listed in abundance order) and most likely to be impacted by the Array (volume 2, chapter 11) are:
- black-legged kittiwake Rissa tridactyla (hereafter referred to as “kittiwake”);
- common guillemot Uria aalge (hereafter referred to as “guillemot”);
- razorbill Alca torda;
- Atlantic puffin Fratercula arctica (hereafter referred to as “puffin”);
- northern fulmar Fulmarus glacialis (hereafter referred to as “fulmar”); and
- northern gannet Morus bassanus (hereafter referred to as “gannet”).
- Seabird species diet and foraging behaviour determine the extent to which individual species are impacted and can respond to changes in prey availability. This is discussed further in section 20.9.10. A summary of their typical feeding strategies and prey species of key seabird species that have the potential to be impacted by the Array have been outlined in Table 20.18 Open ▸ .
Table 20.18: Diet and Feeding Strategies of Key Seabird Species
20.9.5. The Key Prey Species
- The key fish and shellfish prey species likely to be present within the fish and shellfish study area, are the small shoaling forage fish sandeel, herring, mackerel and sprat. Volume 2, chapter 9 identified that these fish species are IEFs. The abundance of each of these species within the fish and shellfish study area and their relative importance to predators is discussed in the species summaries below.
Sandeel
- Sandeels, resembling small eels, predominantly feed on plankton of variable sizes, ranging from small plankton eggs up to larger, energy-rich copepods. In Scottish waters, there are five sandeel species, with the lesser sandeel and greater sandeel Hyperoplus lanceolatus being the most abundant in the vicinity of the site boundary. Lesser sandeel (hereafter referred to as “sandeel”) and Raitt’s sandeel Ammodytes marinus are listed as PMFs and listed as protected features within the Turbot Bank Nature Conservation MPA, which occurs within the fish and shellfish ecology study area.
- As well as being abundant in Scottish waters, sandeels are highly nutritious and therefore serve as the preferred prey item for several fish species, seabirds, seals, dolphins and whales. Sandeel represent an important link between the lower and upper levels of the marine food web, feeding on plankton and preyed upon by marine predators like cod, kittiwake and harbour porpoise (NatureScot, 2022).
- Sandeel have a close association with sandy structures into which they burrow. They are largely stationary after settlement and show a strong preference to specific substrate types. For the purposes of considering sandeel habitat suitability, gravelly sand (between 30% and 5% gravel), slightly gravelly sand (between 5% and 1% gravel) and sand (under 1% gravel) in the European Marine Observation and Data Network (EMODnet) substrate data were classified as preferred habitat and sandy gravel (between 30% and 80% gravel) as marginal habitat. The substrates classified as preferred and marginal habitats all have a sand to mud ratio of nine to one or higher. Where no shading from sunlight is present, the habitat in that area is considered unsuitable for sandeel (volume 3, appendix 9.1).
- As described in volume 2, chapter 9 and volume 3, appendix 9.1, through predicted distribution modelling by Langton et al. (2021) the whole site boundary has extremely low probability of sandeel presence. Abundance data from grab sampling and epibenthic trawls within the site boundary indicated that when sandeel were present, it was predominantly in the north-west section of the site boundary, which aligns with the marginal and preferred habitats for species within this section. Higher densities within the north-west section were found closer to the coasts or towards the Firth of Forth (volume 3, appendix 9.1).
Herring
- Herring is a small shoaling forage fish which is a commercially important pelagic fish. Commonly found across much of the North Sea, herring filter feeds on plankton and minute sea creatures, supplemented by small sprats and fry of other fish species (British Sea Fishing, 2022).
- Nursery grounds for herring are abundant across the east Scottish and Northumberland coastlines (Ellis et al., 2012). These areas serve as feeding grounds for post-larvae juveniles and sub-adults, before individuals reach sexual maturity and migrate further offshore (International Council for the Exploration of the Seas (ICES), 2006).
- Herrings are a key prey species for numerous fish, birds and marine mammals. Herring are listed on the Scottish Biodiversity List (SBL) and as a PMF and are therefore considered a high priority species for conservation actions in Scotland (Fauchald et al., 2011; Casini et al., 2004).
- Spawning for herring usually takes place in shallow areas between approximately 15 m and 40 m depth. Herring utilise specific benthic habitats during spawning (e.g. gravel, shells and small stones) with muddy sediments considered unsuitable due to increased egg mortality via asphyxiation as a result of coarse sediment of these environments blocking the pores of the eggs.
- Herring are considered hearing specialists with an increased sensitivity to underwater noise and are therefore vulnerable to injury or disturbance from activities which generate underwater noise, such as pile driving (volume 3, appendix 10.1).
- North Sea herring fall into a number of different ‘races’ or stocks, each with different spawning grounds, migration routes and nursery grounds (Coull et al., 1998). North Sea autumn-spawning herring have been divided into three, mainly self-contained stocks – the Buchan/Shetlands, Banks/Dogger and Bight/Downs herring groups, which show differences in spawning areas and spawning periods. The Buchan/Shetlands stock spawns off the Scottish and Shetlands coasts in August and September and is the most relevant to the Array as the closest to the site boundary and fish and shellfish ecology study area (volume 3, appendix 9.1).
- Herring spawning grounds are most accurately mapped using a combination of herring larval data and sediment particle size analysis (PSA), as recommended by Boyle and New (2018).
- Site-specific surveys illustrated that the overwhelming majority of (95%) of the site boundary has unsuitable sediment composition for herring spawning. The only four stations considered suitable for herring spawning were sparsely distributed in the north-west and centre of the site boundary (volume 3, appendix 8.1). Preferred habitats are located directly north of the site boundary, in line with spawning grounds detailed in Coull et al. (1998).
Mackerel
- Mackerel is a small, fast, predatory fish closely related to tuna Thunnini sp. which hunt in vast shoals for smaller fish and sandeel.
- As a vital prey species for larger fish, birds and marine mammals, mackerel are listed as PMFs in Scottish waters (NatureScot, 2020).
- Mackerel are migratory fish that are common throughout the UK, typically arriving in spring and early summer when they will feed actively before they migrate to warmer seas for the autumn and winter months for spawning. During this spawning period, their feeding activity diminishes significantly.
- Observations have been made that mackerel are arriving in UK waters earlier and leaving later every year, possibly as a result of rising sea temperatures. This has been linked to the complete absence of mackerel in areas around the south of the UK during the winter months.
- The absence of a swim bladder enables mackerel to swiftly adjust their depth, allowing them to maintain constant movement (British Sea Fishing, 2022).
- As described in volume 3, appendix 9.1, mackerel have low intensity nursery grounds which overlap with the site boundary, with no spawning grounds identified within or in proximity to the site boundary (Ellis et al., 2012). Mackerel spawning behaviour involves the release of eggs into the water column, where fertilisation also occurs (Walsh and Johnstone, 2006), indicating a low level of reliance on sedimentary habitats for spawning. Mackerel spawn over summer months from May to August. The presence of mackerel nursery grounds is not supported by outputs from Aires et al. (2014), with no modelled observations of “0 group fish” on the east coast of Scotland.
Sprat
- Sprat is a small foraging fish which is widespread across the UK, inhabiting water depths ranging from a few metres to approximately 100 m.
- Predominantly feeding on fish eggs, larvae, and plankton, sprat are a major part of the marine food chain in the North Sea. Sprat provide a vital food source for more or less all predatory fish species found in UK waters, as well as seabirds and marine mammals.
- As described in volume 3, appendix 9.1, sprat have an unidentified intensity spawning and nursery grounds over the majority of the site boundary.
20.9.6. How the Food System Works
- The transfer of energy moves up through the trophic levels of the food chain, starting at the bottom level where producers like phytoplankton and algae in the marine environment produce their own food by harnessing energy from the sun through the process of photosynthesis. Primary consumers, such as zooplankton, then feed on the phytoplankton to gain energy which is then transferred up each trophic level of the food chain.
- The marine environment typically follows a ‘wasp-waist’ trophic structure, where mid-trophic level species have lower diversity, compared to high diversity in both high and low trophic levels. These mid-trophic level species are crucial in the functioning of ecosystems (Rice, 1995). The main prey species found within the site boundary are sandeel, herring, mackerel and sprat. These fish link the lowest trophic level (e.g. phytoplankton) to the highest (e.g. marine mammals) (Mackinson and Daskalov, 2007; Feuchald et al., 2011; Lynam et al., 2017).
- Phenology plays an important role in how the food chain operates because many species have evolved elaborate behaviour and life history strategies that favour certain periods of the year for growth and reproduction and minimise the exposure of sensitive life stages in more stressful periods (Rubao et al., 2010). Any changes to phenology as a result of climate change can affect the lowest trophic levels and cause a cascade effect up through the food chain. For example, changes in sandeel populations will have a resulting effect on higher trophic species such as seabirds (Burthe et al., 2012; Lynam et al., 2017). This is further discussed in section 20.9.10.
- Section 20.9.4 describes the key fish, seabird and marine mammal predator species and their typical prey species. It can be noted that whilst the key prey species in section 20.9.5 are components of most predators’ diets, they vary in their importance. For example, kittiwake are more reliant on sandeel than the other key seabird species potentially present within the site boundary. Kittiwake would therefore be more sensitive to changes in sandeel distribution and availability. This is discussed further in section 20.9.10.
20.9.7. Future Ecosystem Baseline
- The EIA Regulations require that a “a description of the relevant aspects of the current state of the environment (baseline scenario) and an outline of the likely evolution thereof without implementation of the Array as far as natural changes from the baseline scenario can be assessed with reasonable effort, on the basis of the availability of environmental information and scientific knowledge” is included within the Array EIA Report.
- If the Array does not come forward, an assessment of the ‘without development’ future baseline conditions has also been carried out and is described within this section.
Climate change effects
- The baseline environment for the physical and biological components of the ecosystem are subject to natural fluctuations over time. These changes will occur with or without the development of the Array due to natural variability. Therefore, it is important that when undertaking assessments of effects, any potential impacts must be considered within the context of the envelope of change that might occur over the timescale of the Array. Rising sea levels, increase sea temperatures and storminess are all likely to alter the future baseline conditions. In terms of physical processes, this is unlikely to have the effect of significantly altering tidal patterns and sediment transport regimes offshore at the site boundary. The return period of the wave climates would be altered (e.g. what is defined as a 1 in 50 year event may become a 1 in 20 year event) as deeper water would allow larger waves to develop. There is, however, uncertainty in the precise impacts climate change will have on prevailing wave climates within the North Sea and beyond.
- Sea surface temperatures (SSTs) around Scotland are strongly influenced by the atmosphere (heat flux) and ocean circulation (advection). Temperature variability in Scottish waters is provided regularly as part of the Scottish Ocean Climate Status Report (Hughes et al., 2018). Hughes et al. 2018 most recent report showed that Scottish waters (coastal and oceanic) have warmed by 0.05°C to 0.07°C per decade, calculate across the period of 1870 to 2016. Figure 20.2 shows an increasing trend in SST from 1893 to 2018 in all Scottish Marine Regions (SMRs) with trends for each region also ranging between 0.05°C and 0.07°C (Marine Scotland, 2024).
- Changes in temperature will affect the biological environment baseline (including benthic and intertidal ecology, fish and shellfish ecology, marine mammals and seabirds). Fish will be affected at all biological levels (cellular, individual, population, species, community and ecosystem) both directly and indirectly. For example, as sea temperatures rise, species adapted to cold water (e.g. herring and cod) will begin to disappear while warm water adapted species will become more established, creating a shift in the baseline. Changes in the stratification of water columns and plankton production may occur as a result of increased spring storms which may cause knock on effects through the food chain (Morison et al., 2019). The various changes that may occur as a result of climate change are uncertain and therefore it is difficult to predict the future baseline scenario with accuracy.
- Changes in ocean temperature, ocean acidification, water stratification and nutrient availability, as a result of climate change, are affecting the abundance and diversity of communities at all trophic levels (Walther, 2010). Effects have been identified over a variety of timescales. Short-term variability in environmental conditions impacts interactions between trophic levels and species (Howells et al., 2017). Limitations in prey availability can adversely affect top predators, with population level changes likely to occur over longer timescales, propagating up trophic levels with prolonged exposure (Frederiksen et al., 2006; Howells et al., 2017).
- The ability of fish species to move in response to temperature varies depending on a range of factors, including their physiological capacity to acclimatise and respond to the change as well as their degree of geographical attachment or how their prey respond. Where a species has a strong geographical attachment, the result can be a localised decline in species (Wright et al., 2020). Effects on prey species are further discussed in section 20.9.8.
- There is increased research into the effect of ocean acidification on fish physiology and early survival (Wright et al., 2020). As stated in paragraph 150, ocean acidification is a consequence of climate change due to chemical processes related to increased temperatures and increasing concentrations of carbon dioxide dissolving in seawater. The resulting decrease in pH is affecting phytoplankton which can inhibit shell generation of calcifying marine organisms and skeletal development of larval fish, with potential consequences for forage species (Riebesall et al., 2013). Impacts as a result of ocean acidification are however difficult to predict as species and population level due to the complexity of the different trophic levels within the food web (Heath et al., 2012).
- Understanding climate change impacts on top predators is fundamental to marine biodiversity conservation, due to their increasingly threatened populations and their importance in marine ecosystems (Olgeret et al., 2022). The long lifespans and large-scale mobility of top predators such as seabirds and marine mammals integrates information from the bottom to the top of the food chain and can serve as a sentinel of ecosystem change (Hazen et al., 2019). Burthe et al. (2014) conducted a long-term monitoring study (36 years) which found that the majority of a wide range of seabird species (including fulmar, kittiwake and puffin) demonstrated a negative response to increased SST in terms of their population size, breeding success and adult survival. Gannets were found to be less vulnerable which could be due to their higher flexibility in foraging behaviour (particularly in terms of exploiting fisheries discards as an alternative food source) although it is more likely due to a lack of data for this species (Burthe et al., 2014). Cliff nesting species, such as kittiwake and razorbill, may be more sensitive to nest failure as a result of high winds and storm surges caused by climate change (Newell et al., 2015).
- Overall, gannet are thought to be buffered from the impacts of climate change, mostly relating to their ability to access a wider variety of prey, but they may be sensitive to controls on fisheries discards (Johnston et al., 2021). Guillemot, kittiwake, puffin and razorbill abundances have been closely linked to the success of their prey, particularly sandeel (Burthe et al., 2014).
- Most marine mammal and climate change related studies to date have focused on effects of sea ice change however new studies are beginning to be published that consider the broader impacts of climate change on marine mammals. The main impacts are geographic range shifts, reduction in suitable habitats, food web alterations and increased prevalence of disease. Increased SSTs and resulting marine mammal range shifts are leading to novel interactions, increased predation risk and competition for species (Waggitt and Evans, 2020; Martin et al., 2023).
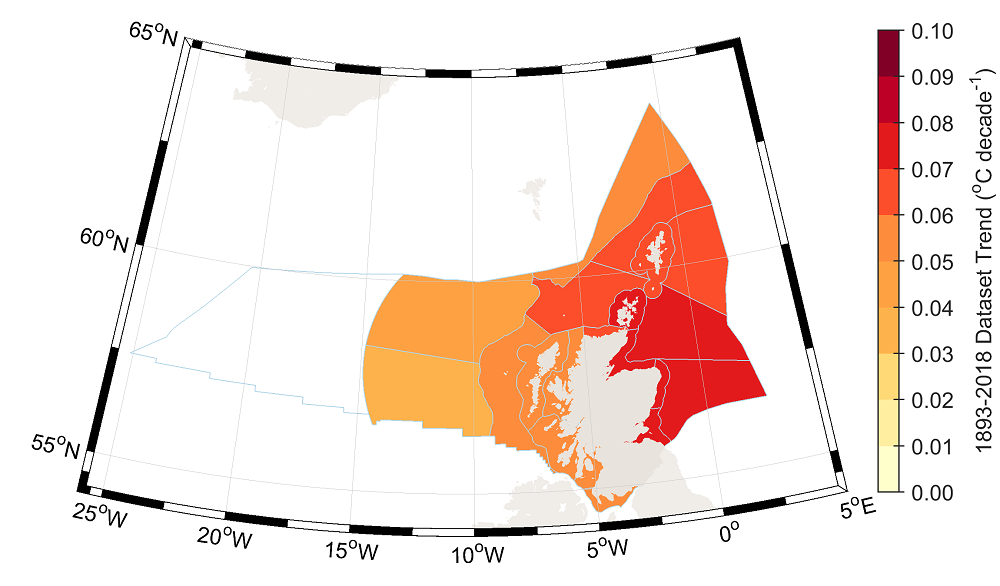
Figure 20.2: Sea Surface Temperature Trend from the ISST Data Product for the Observational Period (1893 to 2018) Averaged by SMRs and Offshore Marine Regions (OMR) (Marine Scotland, 2024)
Highly Pathogenic Avian Influenza (HPAI)
- Seabirds have been severely affected by avian influenza. The most recent HPAI outbreak began in 2021 and has now spread through more than 70 bird species. The virus has moved into species such as guillemot, razorbill, and kittiwake, with dead birds washing up on beaches in Wales, and along the eastern coasts of Scotland and England. At this stage, with little quantitative information, it is difficult to conclude to what extent population levels have been impacted by HPAI. Further information as to how HPAI has been considered in the site-specific surveys of the offshore ornithology study area can be found in volume 3, appendix 11.1.
Sandeel fishery closure
- As described in volume 3, appendix 9.1, the highest density of the sandeel population is focussed on the Wee Bankie (approximately 57 km west of the site boundary), however sandeel do range across much of the North Sea. In the early 1990s, there was a substantial industrial sandeel fishery on the Wee Bankie, Marr Bank and Berwick Bank sandbanks. By 1993, landings from this area had peaked at over 100,000 tonnes (Greenstreet et al., 2010b). In 2000, this industrial sandeel fishery was closed in response to concerns that the fishery was having a deleterious effect on sandeel stocks within the Forth and Tay SMR.
- In 2000, the first year of the closure of the Forth and Tay SMR sandeel fishery, high levels of recruitment, combined with a lack of any significant fishing activity resulted in an immediate and substantial increase in the biomass of sandeel on the Wee Bankie sandbank (Greenstreet et al., 2010b). However, between 2001 and 2010, sandeel biomass steadily declined to levels that were similar to those observed when the sandeel fishery was active (Greenstreet et al., 2010b). This was thought to be due to the absence of sustained recruitment, meaning that predation and other causes of natural mortality still exceeded population growth (Greenstreet et al., 2010b).
- As described in the Array Derogation Case due to the concerns about stock levels, a sandeel fishery ban was implemented in English waters from 2021 through 2023 for UK vessels (Horton, 2022). Following the initiation of a consultation by the Scottish Government in 2023, in January 2024, the Sandeel (Prohibition of Fishing) (Scotland) Order 2024 was established, closing sandeel fishing in Scottish waters from 2024 onwards. The decision considers the role of sandeel in the marine ecosystem(Scottish Government, 2024).
- Additionally, the UK government conducted a public consultation on spatial management measures for sandeel fishing in English waters of the North Sea. This consultation followed a 2021 call for evidence, revealing concerns about the impact of industrial fishing on the marine environment. Expert reports indicated that prohibiting sandeel fishing in the North Sea would benefit seabirds, other fish species, and marine mammals. Over 95% of respondents supported some form of prohibition, with a majority favouring the closure of all English waters. Consequently, the UK government decided to prohibit sandeel fishing within English waters of ICES Area 4 (North Sea) starting from 26 March 2024, before the next sandeel fishing season (Defra, 2024).
20.9.8. Existing Pressures on Prey Species
- Before assessing the potential effects of the Array on prey species at an ecosystem level, it is important to understand the existing pressures on each prey species.
- The North Sea is one of the most anthropogenically impacted marine ecosystems (Halpern et al., 2015; Emeis et al., 2015). Small shoal fish in mid-level trophic levels experience top-down pressure from commercial fisheries whilst bottom-up processes driven by temperature, have dominated changes to planktonic groups since the 1960s. These pressures propagate up and down the food chain, with mid-trophic fish linking the pressures between the upper and lower trophic levels (Lynam et al., 2017).
- Forage fish landings constitute approximately one-third of global landings of marine fish, not including losses from bycatch discards (Alder et al., 2008). Historically, sandeel have been targeted commercially for their oil and use as an animal feed and fertiliser. Despite being managed, the majority of sandeel stocks have experienced severe declines due to a combination of overfishing and the effects of climate change (NatureScot, 2022). In March 2024, the UK government introduced a prohibited sandeel fishing within English waters of ICES Area 4 (North Sea) (Defra, 2024). Further details of this sandeel fishery closure can be found in paragraph 157 to 160.
- As described in volume 3, appendix 9.1, herring are a commercially important pelagic fish in the North Sea which was targeted in the vicinity of the site boundary. The herring stock collapsed entirely in the 1970s as a consequence of overfishing (Scottish Herring, 2023). Since then, stocks have shown signs of recovery supported by a herring recovery plan implemented for the North Sea in 1996 and a ban on discards for pelagic fisheries, including for herring, from 2015. Active management is however still required to avoid a recurrence of the collapse (Dickey-Collas et al., 2010).
- The prey species present in the marine ecosystem within which the Array occurs, are also an important food source for larger fish. For example, plaice, cod, haddock, whiting, saithe, tope shark and spurdog all include prey forage species in their diet such as sandeel, herring, sprat and mackerel. Additionally, diadromous fish species are also likely to feed on these species. Volume 2, chapter 9, identified the following diadromous species are likely to migrate through the fish and shellfish ecology study area: Atlantic salmon, sea trout, sea lamprey, European eel, allis shad and twaite shad.
- As described in section 20.9.7, changes to the baseline environment as a result of climate change will have effects on marine fish species across all trophic levels. In terms of prey species, sandeel and herring are particularly vulnerable to the effects of climate change.
- Sandeel are one of the most important trophic links between plankton and top predators in North Sea ecosystems however climate driven changes to plankton and zooplankton have led to declines in the abundance and nutritional quality of sandeel since 2000 (Macdonald et al., 2015; Clausen et al., 2017; Wanless et al., 2018; MacDonald et al., 2019). This has caused knock-on effects up through the food chain.
- Sandeel are also impacted directly by climate change through their metabolic rate, which can in turn affect the success of their reproduction and increase their mortality rate (MCCIP, 2018; NatureScot, 2022). Increased temperatures have been observed to cause inhibited gonad development in sandeel, which means warmer seas can delay the spawning time and lead to reduced reproductive success (Wright et al., 2017). A key factor in sandeel larval success is synchrony between the larval hatching times and the spring zooplankton bloom. Adult sandeel feed on zooplankton in the spring and summer months; building up lipids to survive the winter period buried in sand when plankton production is lower. Increased temperatures lead to increased energy usage whilst overwintering, meaning less energy can be allocated to gonad development. (Boulcott and Wright, 2008; Wright et al., 2017). The delay to spawning time caused by inhibited gonad development can therefore lead to later larval hatch times, earlier zooplankton blooms and a resulting decrease in zooplankton available for sandeel to feed upon. This consequently can cause a reduction in sandeel growth and survivorship and ultimately low recruitment of the species (Réginer, Gibb and Wright, 2017).
- The life cycle of sandeel ties them to sandy sediments of a particular grain size which they burrow into at night and during the winter months. This means that their ability to move and redistribute to new suitable habitats in response to rising sea temperature relies on larval distribution (Macdonald et al., 2015).
- Herring are also constrained as demersal spawners, by their requirement to spawn at specific locations, depositing their sticky eggs on coarse sand, gravel, small stones and rocks (Wright et al., 2020).
20.9.9. Effects of the Array on Prey Species
- This section assesses the potential effects of the Array on prey species and any impacts on physical processes which may impact prey species indirectly by altering their availability to food sources such as plankton and zooplankton.
- Information to support this assessment has been extracted from the relevant receptor topic Array EIA Report chapters. Conclusions on LSE1 have also been extracted from these chapters. Each assessment of an effect focuses on the prey species most vulnerable to the impact and therefore represents the greatest potential impact.
Potential impacts on prey species
- Volume 2, chapter 9 identified that the following potential impacts as a result of the Array could result in the following potential impacts on fish and shellfish and benthic ecology:
- temporary habitat loss and disturbance;
- long-term habitat loss and disturbance;
- colonisation of hard structures;
- underwater noise impacting fish and shellfish receptors;
- underwater noise from the operation of floating wind turbines and anchor mooring lines impacting fish and shellfish receptors;
- increased SSCs and associated deposition; and
- effects to fish and shellfish receptors due to EMFs from subsea electrical cabling.
- Of the potential impacts, the first two were assessed as having minor adverse effects on marine fish (including prey species) which would not result in a significant change in prey species population. A summary of the assessment of these impacts is provided in the following sub-sections.
- The colonisation of hard structures has the potential to affect numbers of prey species and so is described in more detail, drawing on finds of volume 2, chapters 8, 9 and 10. While not likely to have a significant effect during the operation and maintenance phase, this impact could have an effect in the decommissioning phase if hard structures are left in-situ.
Temporary habitat loss and disturbance
- As discussed in volume 2, chapter 9, in general, mobile fish are able to avoid areas subject to temporal habitat disturbance. Of the key prey species, sandeel and herring are more sensitive to temporary habitat loss as they spawn on or near the seabed however recovery is expected to occur quickly as the sediment recovers post-construction and recolonisation occurs. Furthermore, the conditions in the fish and ecology study area are largely unsuitable for herring and sandeel habitats, and so there is limited disturbance to the spawning of these species. While there is a small overlap with herring spawning grounds, the impact is expected to be very limited due to the context of available favourable sediments habitat outside and across the fish and shellfish ecology study area.
Long term habitat loss and disturbance
- As discussed in volume 2, chapter 9, long term habitat loss and disturbance may arise due to the operation and maintenance phase of the Array. As with the potential impact of temporary habitat loss and disturbance, the most sensitive species are sandeel and herring. They are expected to recover quickly as the sediments recover following installation of Array infrastructure and adults and larvae recolonise the sandy sediments. Sandeel are particularly sensitive to long-term habitat loss and disturbance because of their specific habitat requirements (e.g. sandy sediments) for spawning and burrowing at night and through the winter. However, while sandeel are assessed to have medium sensitivity to this impact, the impact is expected to be limited in extent (particularly in the context of available habitats in the fish and shellfish ecology study area and the wider northern North Sea). Given the limited availability of favourable sediments within the site boundary, significant effects are not predicted.
- Herring habitat is largely unsuitable within the site boundary, and with spawning grounds existing outside the site boundary in the wider fish and shellfish ecology study area. There is, however, a small overlap with the herring spawning habitat and the fish and shellfish ecology study area. However, the area of herring spawning grounds affected by this impact is expected to be very limited (being limited to the site boundary only), in the context of available favourable sediments habitat outside and across the fish and shellfish ecology study area.
Colonisation of hard structures
- Volume 2, chapters 8, 9 and 10 discussed how the introduction of infrastructure within the Array may result in the colonisation of foundations, scour protection and cable protection. Since these hard structures are added to the areas typically characterised by soft, sedimentary environments, the resulting change of habitat type acts like an artificial reef. Anthropogenic structures on the seabed attract many marine organisms including benthic species normally associated with hard biological structures (e.g. blue mussel) (Karlsson et al., 2022). Additionally, man-made structures may also have direct effects on fish through their potential to act as fish aggregation devices (Peterson and Malm, 2006). While the reef effect can affect the existing biological soft sediment communities it can also have potentially beneficial effects on the marine ecosystem.
- The colonisation of new habitats may potentially lead to the introduction of INNS. This may have resulting impacts on benthic and fish and shellfish populations as a result of competition. A study into the spread of INNS by wind farm hard structure colonisation suggested the risk of this occurring was minor, and requires more research to fully understand, with implementation of precautionary built-in measures recommended to prevent spread where possible (Baulaz, et al., 2023). Potential adverse effects of the introduction of INNS are discussed further in volume 2, chapter 8.
- Artificial reefs can act as stepping-stones allowing organisms to colonise areas not typical of their species which can increase the connectivity between natural sub-populations (Coolen et al., 2017). The impacts of this can extend beyond the local scale of a single operation (e.g. the Array) with multiple adjacent offshore wind farms creating stepping stones across wider areas and creating a large-scale effect (Degraer et al., 2020). For example, the Array is close to three offshore wind farms: Seagreen 1 Offshore Wind Farm, Kincardine Offshore Wind Farm and Seagreen 1A Project (volume 2, chapter 15). This cumulative effect does not extend to benthic communities which are unlikely to be suited to the sedimentary habitats between projects and therefore will only colonise the hard structures of individual projects. Despite this, increased vessel presence could provide vectors and stepping stones for larval species and INNS. As species become established on and around the artificial hard structures, they can start producing larvae, with one study demonstrating that networks of oil and gas infrastructure in the North Sea could facilitate ecological connectivity by acting as stepping stones for larval connectivity (Henry et al., 2018) (volume 2, chapter 8). There is some evidence (although with uncertainties) that some fish and shellfish populations are likely to benefit from introduction of hard structures. See further details on the effects of the colonisation of hard structures on benthic and fish and shellfish ecology from paragraph 182 et seq.